Während die Sonne als Energiequelle für Lebensprozesse nahezu unerschöpflich ist, sind die für den Aufbau organischer Substanz lebensnotwendigen chemischen Elemente nur in begrenzter Menge vorhanden. Daher ist das Leben auf der Erde auf die Wiederverwertung essentieller Elemente angewiesen. Durch fortlaufende Auf- und Abbauprozesse zirkulieren die Elemente in den Organismen, Ökosystemen und der Biosphäre. Dieser Prozess wird als Stoffkreislauf bezeichnet. Treibende Kraft für die Stoffkreisläufe ist ein Zusammenwirken von biologischen, geologischen und chemischen Prozessen unter Beteiligung biotischer und abiotischer Komponenten der (Öko-)Systeme. Sie werden daher auch als biogeochemische Stoffkreisläufe bezeichnet (s. CAMPBELL 1997, S. 1254).
An dem lebensnotwendigen "Recycling"-Prozess in den biogeochemischen Stoffkreisläufen sind die Lebewesen durch Nahrungsaufnahme und Stoffwechsel maßgeblich beteiligt. Lebewesen tauschen durch Nährstoffaufnahme, Atmung und Ausscheidung von Abfallprodukten ständig chemische Bestandteile mit der Umwelt aus. Grundsätzlich lassen sich bei den Organismen verschiedene Ernährungstypen und Ernährungsformen unterscheiden, die über die Nahrungskette eine vielfältig vernetzte Ernährungsgemeinschaft entwickeln und sich entsprechend ihrer Nahrungsquelle verschiedenen Trophiestufen (= Nahrungsstufen) zuordnen lassen: Produzenten , Konsumenten und Reduzenten bzw. Destruenten (s. Nahrungskette und Nahrungsnetz ).
Nahrungsgrundlage für alle anderen Organismen sind die Produzenten. Dazu gehören grüne Pflanzen, Algen und einige Bakterien, die in der Lage sind, anorganische Verbindungen in organische körpereigene Verbindungen umzuwandeln. Konsumenten und Destruenten dagegen müssen organische Substanzen aufnehmen, um Energie und lebenswichtige Substanzen für ihren Stoffwechsel zu gewinnen. Die von den Produzenten synthetisierte organische Substanz wird über verschiedene Konsumentenstufen im Nahrungsnetz weitergegeben und schließlich von den Destruenten abgebaut (s. Zersetzung).
Die in der organischen Substanz
gebundenen Elemente gelangen so in pflanzenverwertbarer
Form zurück in die abiotische
Umwelt (Atmosphäre, Boden, Wasser) und stehen den pflanzlichen
(Primär-) Produzenten wieder
für den Aufbau neuer
organischer Substanz zur Verfügung (vgl. CAMPBELL 1997,
S. 1249 und 1254). Damit schließt sich der Nährstoffkreislauf
(s. Abb.).
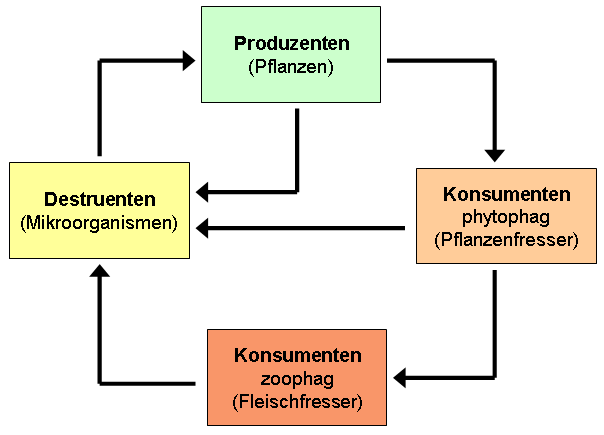 |
| Nährstoffkreislauf und der Nahrungsbeziehungen (vereinfachte Darstellung) |
Für jedes der in einem Lebewesen
vorkommenden Elemente können die spezifischen Stoffkreisläufe
isoliert betrachtet werden. Lebenswichtige Stoffkreisläufe
sind z.B. die der Elemente Kohlenstoff (C) und Sauerstoff
(O), Stickstoff (N), Phosphor (P) und Schwefel (S). Eine
zentrale Rolle für die Organismen spielen dabei der
Kohlenstoffkreislauf und
der Stickstoffkreislauf.
Weitere Informationen:
- Kohlenstoffkreislauf
- Stickstoffkreislauf
Literatur:
CAMPBELL, N.A. (1997): B iologie.
Heidelberg; Berlin; Oxford: Spektrum.
Gisi, U./
Schenker, R./ Schulin, R./ Stadelmann, F.X./ Sticher, H.
(1997): Bodenökologie
- 2. Auflage - Stuttgart; New York: Thieme.
Scheffer, F./ Schachtschabel, P. (2002): Lehrbuch der Bodenkunde
- 15. Auflage -. Heidelberg; Berlin: Spektrum Akademischer
Verlag.
![]()
Kohlenstoffkreislauf
Kohlenstoff ist Grundbaustein aller organischen Verbindungen und zirkuliert über die Nahrungskette im Ökosystem. Als Kohlenstoffquelle dient primär Kohlenstoffdioxid (CO2) aus der Atmosphäre. Stärker als bei anderen Elementen ist der Weg des Kohlenstoffs durch das Ökosystem mit dem Energiefluss gekoppelt und an gegenläufige biologische Prozesse gebunden: Assimilation (Photosynthese) und Dissimilation (Zellatmung). Grundlegend für den Kohlenstoffkreislauf sind die Produktion von CO2 beim Abbau organischer Substanzen (= Zellatmung/ Dissimilation) und der Konsum von CO2 beim Aufbau der organischen Verbindungen (= Photosynthese / Assimilation).
Bei der Assimilation wird Kohlenstoffdioxid (CO2) aus der Atmosphäre aufgenommen und mit Wasser unter Nutzung der Sonnenenergie zu organischen Verbindungen (Kohlenhydrate) aufgebaut. Dieser Prozess wird als Photosynthese bezeichnet. Nur grüne Pflanzen, Algen und Cyanobakterien sind in der Lage, photosynthetisch organische Verbindungen aufzubauen und dabei die Energie des Sonnenlichtes chemisch zu binden. Sie werden daher auch als Produzenten (s.o.) bezeichnet. Die von ihnen photosynthetisch erzeugte organische Substanz dient allen anderen Organismen direkt oder indirekt über die Nahrungskette als Kohlenstoff- und Energiequelle. Gleichzeitig wird bei der Photosynthese Sauerstoff (O2) freigesetzt, der für die Zellatmung benötigt wird.
Bei der Dissimilation werden die organischen Verbindungen abgebaut und die darin gebundene chemische Energie für den Bau- und Betriebsstoffwechsel genutzt. Dabei wird Sauerstoff (O2) verbraucht und das bei der Assimilation gebundene CO2 (wieder) freigesetzt. Dieser Prozess wird als (Zell-) Atmung bezeichnet und ist grundlegend für die Existenz aller Lebewesen, für Konsumenten und Reduzenten , aber auch die Produzenten (s. Abb.).
| Kohlenstoffkreislauf |
Die Rückfuhr von Kohlenstoff aus Dissimilationsprozessen im Boden (s. Bodenatmung) erfolgt ebenfalls überwiegend in Form von CO2 und nur in stark sauerstoffunterversorgten Böden in Form von Methan (CH4). Beide Gase werden über die Bodenoberfläche an die Atmosphäre abgegeben.
Kohlenstoffdioxid (CO2) ist in der Atmosphäre relativ gering konzentriert (ca. 0,03 Vol.-%). Die Umsatzrate für Kohlenstoff bzw. CO2 ist aufgrund der Photosyntheseaktivität der Produzenten aber relativ hoch. Etwa ein Siebtel des gesamten CO2 -Vorrates in der Atmosphäre wird pro Jahr von der Vegetation assimiliert. Diese Menge wird durch Dissimilationsprozesse bei der Zellatmung aller Organismen weitgehend ersetzt, so dass der CO2 -Gehalt in der Atmosphäre durch den Kohlenstoffkreislauf nur geringfügig schwankt.
Durch die Verbrennung fossiler Brennstoffe
wie Kohle, Erdgas und Erdöl werden aber relativ große
Mengen an CO2 zusätzlich freigesetzt. Die
daraus resultierende Erhöhung des CO2-Gehaltes
der Atmosphäre lässt
sich in ihren Folgewirkungen bislang noch nicht endgültig
einschätzen. Relativ gut erforscht sind jedoch die negativen
Auswirkungen menschlichen Handelns auf die globale Klimaentwicklung,
die als sogenannter Treibhauseffekt diskutiert
werden. Auf der anderen Seite fördern höhere CO2
-Gehalte der Atmosphäre aber auch das Pflanzen- und
Algenwachstum.
Weitere Informationen:
- Stoffkreisläufe
- Stickstoffkreislauf
Literatur:
CAMPBELL, N.A. (1997): B iologie.
Heidelberg; Berlin; Oxford: Spektrum.
Gisi, U./
Schenker, R./ Schulin, R./ Stadelmann, F.X./ Sticher, H.
(1997): Bodenökologie
- 2. Auflage - Stuttgart; New York: Thieme.
Scheffer, F./
Schachtschabel, P. (2002): Lehrbuch der Bodenkunde - 15.
Auflage -. Heidelberg; Berlin: Spektrum Akademischer Verlag.
Schroeder, D. (1992): Bodenkunde in Stichworten
- 5. Auflage - Berlin; Stuttgart: Borntraeger.
![]()
Stickstoffkreislauf
Als Strukturbestandteil von Tier- und Pflanzenzellen ist Stickstoff (N) ein (weiteres) Schlüsselelement für Lebensprozesse. So enthalten alle Aminosäuren, Proteine und andere Zellbestandteile wie z.B. Chromosomen (DNA), Chlorophyll und Vitamine Stickstoff als Baustein.
Die permanente Bewegung, Veränderung und Umlagerung des Bioelementes Stickstoff in seinen verschiedenen Formen im Systemkomplex Atmosphäre-Biomasse-Boden wird als Stickstoffkreislauf bezeichnet. Dieser vollzieht sich im Wesentlichen zwischen drei Speichern: dem atmosphärischen Luftstickstoff-Reservoir (ca. 78 Vol.-% N 2 in der Luft), dem in der Biomasse der Lebewesen fixierten Stickstoff und dem in der organischen Substanz des Bodens (Humus) gebundenen Stickstoff. Zwischen diesen Fraktionen erfolgt durch chemische Umsetzungen und Stoffwechselprozesse ein mehr oder weniger intensiver Stickstofftransport und -austausch (LESER 1994, S. 522).
Hinsichtlich der N-Zufuhr in die Ökosysteme lassen sich zwei natürliche Wege unterscheiden: die atmosphärische Deposition und die biogene Stickstoff-Fixierung. Bei der atmosphärischen Deposition gelangen die beiden für Pflanzen direkt verwertbaren Formen des Stickstoffs -Ammonium (NH4+) und Nitrat (NO3-) - im Regen gelöst oder durch Feinstaubablagerungen in den Boden. Hierdurch werden aber nur etwa 5-10 % des verwertbaren Stickstoffs in das System eingetragen (s. Abb.).
Eine weitere N-Quelle ist die biogene Stickstoff-Fixierung. Dabei wird atomarer Luftstickstoff aus der Atmosphäre von einigen Bakterienarten fixiert und zu Ammoniak (NH3) umgewandelt, aus dem organische Stickstoffverbindungen wie z.B. Aminosäuren synthetisiert werden können. Nicht fixierter Ammoniak wird freigesetzt und für andere Organismen verfügbar (s. Abb.). Dieser Prozess ist zwar eine wichtige Ressource für den Stickstoffeintrag ins Ökosystem, spielt aber mengenmäßig im N-Kreislauf eine relativ geringe Rolle. So können freilebende Bodenbakterien der Gattung Azotobacter z.B. maximal 30 kg N pro Hektar und Jahr aus der Atmosphäre fixieren, symbiontisch lebende Bodenbakterien der Gattung Rhizobium z.B. maximal 300-600 kg N/ha/J (vgl. GISI et al. 1997, S. 195).
Eine weitere, nicht natürliche, aber mengenmäßig z.T. sehr bedeutsame Stickstoffquelle stellt die Einbringung von organischen Wirtschaftsdüngern (Gülle und Mist) und anorganischen Kunstdüngern in landwirtschaftlich genutzten Gebieten dar.
Unmittelbares Produkt der Stickstoff-Fixierung und Ausgangspunkt für den biologischen N-Kreislauf ist Ammoniak (NH3), der in der meist schwach sauren Bodenlösung (s. Bodenreaktion) mit den Wasserstoff (H+)-Ionen zu Ammonium (NH4+)-Ionen reagiert, das von den Pflanzen als Primärproduzenten (s. Kohlenstoffkreislauf) direkt als N-Quelle genutzt werden kann (s.Abb.).
Bei ausreichender Sauerstoffversorgung wird der größte Teil des im Boden vorhandenen Ammoniums aber von aeroben Bakterien (z.B. Nitrosomonas und Nitrobacter ) als Energiequelle für Stoffwechselprozesse genutzt. Dabei wird Ammonium zunächst zu Nitrit (NO2-) und dann zu Nitrat (NO3-) oxidiert (s. Abb.). Dieser Prozess verläuft über zwei Stufen und wird als Nitrifikation bezeichnet:
| (1) | NH4+ + 1.5 O2 —> NO2- + H2O + 2H+ |
- 276 kJ mol-1 |
| (2) | NO2-+ 0.5 O2 —> NO3- | - 73 kJ mol-1 |
Optimale Bedingungen für Nitrifikationsprozesse sind: ausreichende Sauerstoffversorgung, neutrale bis schwach saure Bodenreaktion, mittlere Bodenwassergehalte und Temperaturen ziwschen 20 und 25 °C. Nur bei stark saurer Bodenreaktion und Sauerstoffmangel wird der Nitrifikationsprozess so gehemmt, dass im Boden größere Mengen von Ammonium angereichert werden. Bei Sauerstoffmangel können einige Bodenbakterien (z.B. Bacillus oder Pseudomonas ) auch das Nitrat im Boden reduzieren und als Sauerstoffquelle nutzen. Dieser Vorgang wird als Nitratatmung oder Denitrifikation bezeichnet (2 NO3- —> N2 + 3 O2). Das dabei entstehende Stickstoffgas entweicht in die Atmosphäre (vgl. CAMPBELL 1997, S. 1258 und GISI et al. 1997, S. 195/196).
Das bei der Nitrifikation freigesetzte Nitrat wird als Nährsalz über die Wurzel aus der Bodenlösung aufgenommen und zur Synthese organischer Stickstoffverbindungen (z.B. Aminosäuren und Proteine) genutzt. Diese von den pflanzlichen Produzenten synthetisierten organischen N-Verbindungen sind Lebensgrundlage und die einzig nutzbare N-Quelle für alle anderen Organismen (Konsumenten und Destruenten; s. Abb.).
Am Ende der Nahrungskette oder nach Absterben der Organismen werden die in der organischen Substanz gebundenen Stickstoffverbindungen von den Destruenten wieder zu Ammonium (NH4+) abgebaut. Über diesen als Ammonifikation bezeichneten Stickstoffmineralisierungsprozess gelangen große Mengen an Stickstoff zurück in den Boden und können von dort aus wieder aufgenommen oder umgesetzt werden (s. Abb.). Optimale Bedingungen für den Ammonifikationsprozess sind: mittlere Bodenwassergehalte und relativ hohe Temperaturen (bis etwa 50°C).
| Stickstoffkreislauf |
Stickstoff ist zu über 99% in organischen Verbindungen fixiert. Das mengenmäßig bedeutsamste Reservoir für den Stickstoffkreislauf stellt die organische Substanz im Boden (Humus) dar. Je nach Humusart liegt das C/N-Verhältnis , d.h. das Verhältnis von Kohlenstoff zu Stickstoff im Boden in der Regel zwischen 8 und 30. Je kleiner die Zahl, desto enger ist das C/N-Verhältnis und umso besser die Stickstoffverfügbarkeit.
Insgesamt gesehen ist der Stickstoff-Kreislauf
primär
biogener Natur, da in den meisten Ökosystemen der größte
Anteil der zirkulierenden Stickstoffmengen durch Zersetzung der
organischen Substanz und Aufnahme der dabei freigesetzten
Grundbausteine durch die Organismen unmittelbar wiederverwertet
wird (vgl. CAMPBELL 1997, S. 1258 und GISI et al. 1997, S.
194 ff.).
Weitere Informationen:
- Stoffkreisläufe
- Kohlenstoffkreislauf
- Nährstoffhaushalt
Literatur:
Campbell, N.A. (1997): B iologie.
Heidelberg; Berlin; Oxford: Spektrum.
Gisi, U./
Schenker, R./ Schulin, R./ Stadelmann, F.X./ Sticher, H.
(1997): Bodenökologie
- 2. Auflage - Stuttgart; New York: Thieme.
Leser, H.
(Hrsg.) (1994): Westermann-Lexikon Ökologie & Umwelt.
Braunschweig: Westermann.
Scheffer, F./ Schachtschabel, P.
(2002): Lehrbuch der Bodenkunde - 15. Auflage -. Heidelberg;
Berlin: Spektrum Akademischer Verlag.